El VIH nunca ha sido aislado ni purificado de una manera científicamente aceptable.
CONTENIDO
1. Varias declaraciones falsas respecto a las pruebas para VIH por Gallo, Geffen, Gonsalves, et al. (Gallo et al 2006).
2. Las compañías farmacéuticas reconocen que las pruebas para VIH no son específicas para diagnosticar la infección por VIH.
3. El VIH nunca ha sido aislado ni purificado de una manera científicamente aceptable.
4. Las llamadas proteínas del VIH no son marcadores específicos del VIH.
5. El llamado ARN del VIH no es un marcador específico del VIH.
6. Reacciones falsas positivas en las pruebas para VIH .
7. El verdadero significado de ser "VIH-positivo" o "seropositivo".
8. Experimentos propuestos durante el Panel de los Asesores Presidenciales del SIDA en Suráfrica.
9. Conclusiones y recomendaciones.
1.
Varias declaraciones falsas respecto a las pruebas para VIH por Gallo, Geffen, Gonsalves, et al (Gallo et al 2006).
El 4 de marzo de 2006, Robert Gallo junto con activistas pro-antirretrovirales de la “Treatment Action Campaign” en Suráfrica, la “lgtb Men's Health Crisis” en los Estados Unidos, la “Elizabeth Glaser Pediatric AIDS Foundation”, también en los Estados Unidos, y otras (Gallo et al 2006), hizo pública una presunta refutación de un artículo de Celia Farber publicado en el número de marzo de 2006, en Harper's Magazine: "Fuera de control: El SIDA y la corrupción de la ciencia médica" (Farber 2006).
Con respecto a las pruebas para VIH, Gallo y sus co-autores afirman que (Gallo et al 2006):
"Las pruebas para VIH fueron altamente precisas desde que fueron desarrolladas en 1984 y han llegado a serlo aún más a lo largo del tiempo, según que la tecnología subyacente haya evolucionado. Las pruebas para VIH están entre las más precisas disponibles en la medicina actual".
"La prueba de la PCR para detectar la presencia del bichito puede también determinar con precisión el estado de VIH en niños"
"Un diagnóstico de SIDA no puede ser considerado definitivo sin una prueba para VIH".
"Los comentarios de Farber acerca de viajar en avión de Uganda a Australia para cambiar el diagnóstico VIH es simplemente una exageración estulta".
"El riesgo de un falso positivo en la prueba para VIH en África, como en cualquier otro lugar del mundo, es muy pequeño si se sigue el protocolo correctamente. Algunas pruebas de anticuerpos para VIH han sido probadas en África y se las ha encontrado muy precisas. Son las que se usan generalmente. Por ejemplo, la prueba rápida ´Abbott Determine´ usada ampliamente en Suráfrica tiene una especificidad de al menos 98% (y en algunos estudios ha alcanzado prácticamente el 100%). Cuando esta prueba se combina con una segunda ´prueba rápida´ o con una prueba de ELISA para determinar un estado de VIH, el riesgo de un falso positivo es insignificante. La contribución de la tuberculosis y la malaria a los falsos positivos en las pruebas de hoy en día es igualmente insignificante".
"Una aplicación apropiada del protocolo de las pruebas para VIH (lo que incluye realizar por lo menos dos pruebas para VIH) tiene muy pocas posibilidades de dar falso positivo, independientemente del estado de embarzo"
Sin embargo, los datos científicos disponibles no validan estas declaraciones. Varios hechos establecidos científicamente apoyan la aseveración de que las pruebas para VIH no pueden diagnosticar la infección por VIH. A continuación algunos de estos hechos:
2. Las compañías farmacéuticas reconocen que las pruebas para VIH no son específicas para diagnosticar la infección por VIH.
Las pruebas principales para el diagnóstico de infección por VIH son dos pruebas de anticuerpos, ELISA y Western blot, y una prueba genética, la PCR o prueba de "Carga Viral".
Sin embargo, las pruebas de ELISA y Western blot solamente detectan anticuerpos contra lo que se acepta erróneamente ser proteínas o antígenos del VIH. Similarmente, la PCR o prueba de “Carga Viral" solamente detecta copias de fragmentos de ARN que han sido arbitrariamente considerados como el ácido nucleico del VIH. Ninguna de esas pruebas detecta el bichito VIH ni a partículas de VIH.
Las compañias farmacéuticas que producen y comercializan esas pruebas reconocen la imprecisión de las mismas. Esto explica las aparentemente sorpresivas declaraciones incluídas en los instructivos que vienen con los reactivos: "La prueba de ELISA sola no puede ser usada para diagnosticar el SIDA, incluso si varias pruebas de la misma muestra de sangre resultan reactivas y sugieran con alta probabilidad la presencia de anticuerpos anti HIV-1" (Abbott 1997).
Los instructivos para una de las pruebas para administrar el Western blot advierten: "No use esta prueba como la única base para el diagnóstico de la infección por VIH-1" (Epitope Organon Teknika).
En forma similar, el instructivo que acompaña a los reactivos de una prueba frecuentemente usada para la PCR o "Carga Viral" advierte: "la prueba de ampliación genética para monitorizar al VIH-1 no está prevista para ser usada como una prueba rastreadora del VIH ni como prueba diagnóstica para confirmar la presencia de infección por VIH" (Roche 2003).
Por tanto, las compañías farmacéuticas fabricantes de los reactivos para estas pruebas reconocen el hecho de que ni la prueba de ELISA, ni la de Western blot, ni la de "Carga Viral" para VIH son específicas para diagnosticar la infección por VIH.
Es interesante que el único método válido para establecer la sensibilidad y la especificidad de una prueba de laboratorio clínico es comparar la prueba en cuestión con su prueba"estándar de oro". La única prueba "estándar de oro" posible para las pruebas de VIH es el mismo bichito de la inmunodeficiencia humana, VIH. Puesto que el VIH nunca ha sido aislado ni purificado como una partícula viral libre e independiente, tampoco es posible definir correctamente la sensibilidad ni la especificidad de ninguna de estas pruebas. Actualmente, la sensibilidad y la especificidad de las pruebas para VIH, son definidas arbitrariamente, no por comparación con el propio VIH, sino por comparación de las pruebas en cuestión con las manifestaciones clínicas del SIDA, o con el recuento de células T4. Esto explica por que Abbott advierte claramente: "En la actualidad no hay estándar reconocido para establecer la presencia o ausencia de anticuerpos anti VIH-1 en la sangre humana. Por tanto la sensibilidad se calcula basada en los diagnósticos clínicos de SIDA y la especificidad basada en donantes aleatorios" (Abbott 1997).
Puesto que no hay estándar de oro para definir la especificidad de las pruebas usadas para el diagnóstico de la infección por VIH, todos los resultados VIH-positivos deben ser considerados resultados falsos positivos. Además, por todo lo anterior, no es posible identificar a ningún individuo ni como VIH-positivo ni como VIH-negativo.
La gran mayoría de los investigadores del SIDA, periodistas, gente del común y trabajadores de la salud no saben de las limitaciones de estas pruebas porque no tienen acceso a la información pertinente. Adicionalmente, no se da información sobre estos hechos a los médicos y mucho menos al público en general, por parte de las facultades de medicina e instituciones de investigación.
3. El VIH nunca ha sido aislado ni purificado de una manera científicamente aceptable.
Los procedimientos adecuados para el aislamiento y purificación de retrovirus (anteriormente conocidos como bichito tumorales ARN) fueron establecidos desde 1964 (O'Connor et al 1964; De Harven 1065a,b, 1974).
Las fuentes más comunes de material del cual retrovirus pueden ser aislados y purificados son sangre (viremia), tejidos homogeneizados, y el fluído sobrenadante de cultivos de células infectadas (de Harven 1965a,b).
La técnica más frecuentemente usada para el aislamiento y purificación de retrovirus incluye los siguientes pasos principales. (1) Concentración de las partículas virales por centrifugación; (2) Monitorización mediante microscopía electrónica de las partículas virales concentradas; (3) Análisis bioquímico y genético de las partículas virales purificadas; (4) Control de los experimentos para evitar malinterpretar retrovirus endógenos con retrovirus exógenos infecciosos; y (5) Pruebas biológicas para establecer si el retrovirus aislado es en efecto potencialmente patogénico y virulento (O'Connor et al 1964; De Harven 1965a,b, 1974).
Sin embargo, ni Montagnier, ni Gallo, ni Levy et al. cumplieron con esas técnicas cuando anunciaron haber aislado "el bichito del SIDA" en 1983 y 1984 (Barré-Sinousi et al 1983; Papovic et al 1984; Gallo et al 1984; Levy et al 1984). Los primeros dos pasos fueron omitidos; no proporcionaron la evidencia con microscopio electrónico de que el sobrenadante del cultivo "infectado", en la sedimentación de 1.16 gm/ml de sacarosa, estuviera compuesto mayormente por partículas virales concentradas. En cambio, proporcionaron fotografías de microscopio electrónico de linfocitos de los cultivos estimulados y activados, que liberaban partículas similares a retrovirus. Estas mismas partículas, sin embargo, pueden ser vistas en linfocitos de cultivos estimulados y activados pero "no infectados" (Dourmashkin et al 1993).
Desafortunadamente, los experimentos no fueron controlados adecuadamente; dónde está la fotografía de microscopio electrónico del sobrenadante de los cultivos "infectados" y de los "no infectados" sedimentado a 1.16 gm/ml de sacarosa; microfotografías requeridas para determinar si existían o no partículas virales concentradas en ese gradiente de densidad? Y si ellas estaban sólo en los cultivos “infectados”. Adicionalmente, dónde están las fotografías de microscopio electrónico de linfocitos "no infectados" cultivados en idénticas condiciones? Porqué mostrar sólo las de linfocitos “infectados”?
La presunta existencia del VIH fue afirmada después del estudio de proteínas, de la transcriptasa inversa (TI) y de fragmentos de ARN que fueron encontrados en sobrenadante de cultivo, pero que no fueron extraídas directamente de partículas virales purificadas.
Sorprendentemente, la existencia del VIH fue reivindicada indirectamente, sobre la base de la presencia en cultivo de células muy complejos y/o en individuos "VIH-positivos" de (1) proteínas/glycoproteínas tales como gp160/150, gp120, gp41/45/40, p34/32, p24, y p18/17, cada una de las cuales fué anunciada como perteneciente al VIH; (2) enzimas tales como la transcriptasa inversa que supuestamente pertenece al VIH; (3) fragmentos de ARN o ADN que supuestamente pertenecen al VIH (Papadopulos-Eleopulos et al 1993, 1996, 1997a, 1997b, 1997/8; Turner 1996, 1997(1998, 1998; Philptt 1997; Giraldo et al 1999; de Harven 1997/8, 1998, 2002a,b).
Sin embargo, ninguna de esas sustancias ha sido probado que pertenezcan al VIH. Cómo podría probarse que las moléculas encontradas en esos cultivos pertenecen realmente a partículas virales que nunca han sido adecuadamente purificadas? Cómo sería posible demostrar que esas sustancias no son simplemente microvesículas celulares o restos celulares contenidos en los cultivos y que sedimentan en la misma densidad que los retrovirus?. Para probar que esas moléculas, presuntamente consideradas como "marcadores", son parte de un retrovirus llamado VIH, tendría que haber sido absolutamente necesario primeramente purificar las partículas retrovirales, separándolas de todo lo demás.
Sin embargo, mucho tiempo antes de la aparición de los primeros casos de SIDA, los investigadores que trabajaban con "bichito tumorales ARN", actualmente conocidos como retrovirus, sabían claramente que el primer prerrequisito para el estudio de los componentes o moléculas de bichito es obtener preparados de bichito altamente purificados (de-The & O'Connor 1966). Después de la purificación del "bichito de la leucemia murina" por ejemplo, estos autores fueron capaces de emplear métodos químicos especiales (por ejemplo: tween-ether, ribonucleasa, detergentes) para romper las partículas purificadas y extraer los componentes internos (de Thé & O'Connor 1996). Esto nunca se ha hecho con el VIH.
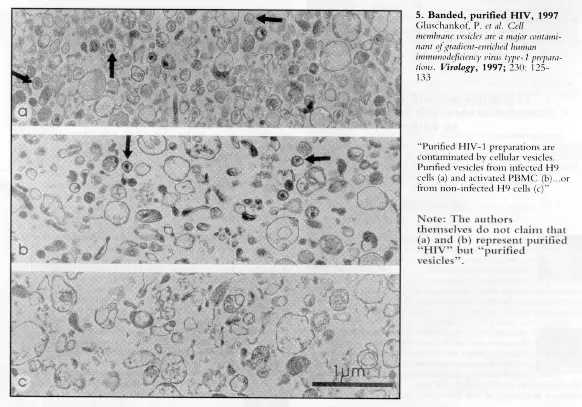
Uno de nosotros ha insistido en que: "La especificidad de los marcadores virales depende del éxito en el aislamiento y purificación. Sin la completa demostración del éxito en el aislamiento y purificación, la identificación de los marcadores virales es extremadamente arriesgada y puede llevar a graves malinterpretaciones de los datos clínicos. Una dramática ilustración de esto se encuentra en la actual investigación del VIH. En este caso, el bichito (VIH) nunca ha sido correctamente aislado, ya que la banda de sedimentación en gradientes de sacarosa en la densidad de 1.16 gm/ml fue erróneamente considerada como de contener solamente bichito, ignorando que el material que sedimenta en esa densidad contiene grandes cantidades de restos celulares y microvesículas celulares (Gluschankof et al 1997; Bess et al 1997). Por tanto, las proteínas y los ácidos nucleicos encontrados en tales bandas a 1.16 muy probablemente son de origen celular y no pueden usarse como marcadores virales. Esta defectuosa metodología ha tenido consecuencias extremadamente serias, como ocurre con las pruebas de anticuerpos anti VIH, ELISA y Western blot, que, se usan mundialmente y peligrosamente, pués carecen de especificidad, como demostraron en 1993 Papadopulos y su grupo (1993), en Australia" (de Harven 1999).
"Más inquietante es el hecho de que algunos ´marcadores virales´ se buscan en material que sedimenta a 1.16, que es la densidad donde se espera sedimenten viriones intactos, pero no sus fragmentos moleculares. Si hubiesen sido disueltas las partículas retrovirales y estas liberaran marcadores moleculares, las muestras a 1.16 permitirían a los investigadores, al menos inicialmente, demostrar por microscopio electrónico partículas virales íntegras. Sin embargo, después de 15 años de la más intensiva búsqueda del VIH, dos grupos independientes finalmente decidieron explorar con microscopio electrónico las características estructurales del material que sedimenta en el gradiente 1.16. Trabajando con sobrenadantes de cultivos de "células T infectadas con VIH-1", ambos grupos hallaron que el material semintando en esa densidad contiene ante todo restos celulares y vesículas de membrana celular, que no pueden ser identificadas como partículas de VIH ni siquiera como objetos similares a bichito" (Gluschankof ete al 1997; Bess et al 1997). Todavía este es el tipo de muestra en el cual los ´marcadores virales´ son identificados en la actualidad y usados para medir los efectos de medicamentos antiretroovirales en ensayos clínicos actuales" (de Harven 1998).
La actividad de la transcriptasa inversa (TI) encontrada en el sobrenadande de cultivos por los investigadores que reivindican haber aislado "el bichito del SIDA" (Barré-Sinousi et al 1983; Papovic et al 1984; Gallo et al 1984; Levy et al 1984) podría también tener un origen celular, puesto que este enzima es ubicua (Ross et al 1971; Beljanski 1972; Varmus 1987; Coffin et al 1997). La TI no es una característica única de los retrovirus, como erróneamente pensaban el grupo de Montagnier, Gallo y Levi.
El VIH tampoco ha sido nunca aislado o purificado como partículas virales intactas. Por tanto, no hay datos científicos que validen la idea de que lo que se conoce como VIH, sea de hecho un bichito!
No existe un solo tubo de ensayo en ningún laboratorio de ninguna parte que contenga partículas purificadas de VIH. Los investigadores que trabajan con lo que ellos creen que es VIH en laboratorios de todo el mundo, es muy probable que no estén trabajando con partículas de VIH. Muy probablemente ellos trabajan con proteínas, enzimas o fragmentos de ARN que han sido arbitrariamente considerados como pertenecientes al VIH.
El hecho de que, después de 25 años de intensa investigación, el VIH no haya sido aislado ni purificado en los términos indicados por la virología clásica, nos indica que la visión del SIDA como una enfermedad viral contagiosa está basada en un microbio que aparentemente no existe!
4. Las llamadas proteínas del VIH no son marcadores específicos del VIH.
A comienzos de los años ochenta, frustrados retrovirólogos que investigaban sobre el cáncer, intentaron probar que el SIDA era una enfermedad retroviral, lo cual fue arbitrariamente decidido basados en lo que ellos erróneamente llamaron "las proteínas del bichito del SIDA", "los enzimas del bichito del SIDA" y "el ARN del bichito del SIDA", los cuales fueron hayados en los sobrenadantes de cultivos, sin haber aislado ni purificado previamente las partículas retrovirales, es decir, sin haberlas separado de microvesículas y restos celulares, como se explicó en la sesión anterior.
El grupo de Montagnier del Instituto Pasteur de Francia, por ejemplo, determinó lo que ellos llaman "antígenos virales" (proteínas virales) a través de una serie de experimentos de inmunoprecipitación (Western blot) utilizando linfocitos de sangre de cordón umbilical en sistemas de cultivos celulares muy complejos: con bichito del paciente 1 como fuente de "antígenos virales", con suero con anticuerpos anti la P24 del HTLV-1 y con suero de los pacientes 1 y 2, y arbitrariamente decidieron que: "habian visto tres proteínas principales: la p25 y proteínas con peso molecular de 80,000 y 45,000. La 45K puede ser debida a la contaminación del bichito con actina celular que estaba presente en los precipitados inmunes de todos los extractos celulares" (Barré-Sinoussi et al 1983). Sin haber purificado previamente las partículas virales, concluyen erróneamente que, "estos resultados, junto con los de inmuno- precipitacion indican que el retrovirus del paciente 1 contiene una proteína principal p25, del tipo similar a la del HTLV-1 pero inmunológicamente diferente" (Barré-Sinoussi et al 1983).
El grupo de Gallo del Instituto Nacional del Cáncer realizó pruebas de Western blot usando "lisados de clones celulares productores de HTLV-III" y suero diluído a 1:500, y, también sin haber purificado previamente las partículas virales, arbitrariamente decidió que, "los antígenos expresados después de la infección viral y reconocidos por el suero humano, incluyeron a p65, p55, p41, p39 y p24. También se detectó una proteína grande con peso molecular de aproximadamente 130,000 y una proteína de 48,000" (Schüpbach et al 1984). Sin embargo, ellos también concluyen que: "estos resultados muestran claramente que los antígenos detectados después de la infección viral pueden corresponder a proteínas de codificacion viral o a antígenos celulares inducidos por la infección" (Schüpbac et al 1984). Adicionalmente, concluyeron que, "se produjo una acumulación grande de p24 y p41, lo cual muestra que estas moléculas son los mayores componentes de la preparacion viral. Por lo tanto la p24 y la p41 fueron, consideradas proteínas estructurales del bichito" (Schüpbach et al 1984).
El grupo de investigadores de Levy, de la Universidad de California en San Francisco, realizó procedimientos de inmuno-fluorescencia indirecta convencional usando "células infectadas" y suero diluído a 1:10. Encontraron anticuerpos contra los que se suponían era ARV (bichito Relacionado con el Sida) en un 88% de SIDA con sarcoma de Kaposi, en un 100% de SIDA con enfermedades oportunistas, en 93% de hombres parceros sensuales de pacientes de SIDA, y en 57% de hombres gayses clínicamente sanos (Levy et al 1984).
Estos tres grupos de investigadores decidieron, arbitrariamente, que las proteínas que hallaron en cultivos de células aparentemente infectados con "el bichito del SIDA" eran "proteínas del VIH". Estas proteínas no habían sido y nunca han sido extraídas directamente de partículas virales aisladas y purificadas. Ellas podrían, por tanto, perfectamente, tener un origen en las células humanas cultivadas.
De otro lado, en 1997, el grupo de Gluschankof en Francia y Alemania, así como el grupo de Bess en los Estados Unidos, demostraron que cuando se sigue el procedimiento rutinario para aislar retrovirus de cultivos que están supuestamente infectados con VIH, no es posible aislar o purificar partículas virales, separadas de microvesículas celulares y de restos celulares, ni siquiera estudiando las bandas donde se sabe que sedimentan retrovirus (Gluschankof et al 1997; Bess et al 1997). Estos investigadores advierten correctamente que, "se debe ser prudente, por tanto, en lo que respecta a la presencia de vesículas celulares cuando se intenten extraer inmunógenos virales (proteínas) de gradientes de densidad" (Gluschankof et al 1997), porque "se han encontrado antígenos celulares humanos en preparaciones de VIH-1" (Gluschankof et al 1997). Por tanto, estos documentos de 1997 de los grupos de Gluschankof y Bess proporcionan una demostración objetiva de que lo que comúnmente llaman "proteínas del VIH" o "antígenos del VIH" o "inmunógenos del VIH" no son marcadores específicos del VIH y podrían muy bien originarse en las células de los cultivos.
A este respecto, nuestros colegas de Perth, Australia, han explicado varias veces que los antígenos, proteínas, glicoproteínas o bandas del Western blot - p120, p41, p32, p24/25, p17/18 – que supuestamente son considerados proteínas específicas del VIH pueden perfectamente no ser codificados por el genoma del VIH, y pueden, de hecho, corresponder a proteínas celulares originadas en las células humanas usadas en los cultivos celulares (Papadopulos-Eleopulos et al 1993, 1997a; Turner 1996, 1997/1998). El componente celular normal actina probablemente corresponda a lo que se conoce como gp41, mientras que la gp120/160 probablemente represente oligómeros de la gp41 (Papadopulos-Eleopulos et al 1993).
Por lo tanto, nadie ha presentado, hasta la fecha, evidencia de que las llamadas proteínas o antígenos del VIH (gp160/150, gp120, gp41/45/40; p34/32, p24, p18/17), sean realmente constituyentes del VIH (Papadopulos-Eleopulos et al 1993, 1996; de Harven 1998, 2002a, 2003; Giraldo 2002a; Giraldo et al 1999).
De otro lado, las proteínas y glicoproteínas enumeradas arriba (conocidas como "antígenos de VIH") se sabe que sólo aparecen cuando uno co-cultiva sangre supuestamente infectada junto con células besugos de pacientes leucémicos, o de linfocitos de cordón umbilical (Papadopulos-Eleopulos et al 1996; de Harven 1998). Muy probablemente, las mismas moléculas se obtendrían de cultivos similares en ausencia de “infección por VIH". Sin embargo, nunca se realizaron estos controles de importancia crucial en los experimentos para aislar al “bichito del SIDA” (de Harven 1998, 2003, 2004), especialmente cuando los investigadores usaron sangre de linfocitos de cordón umbilical. Se sabe que estas células procedentes de la placenta contienen retrovirus endógenos probablemente defectuosos (Panem 1979; de Harven 2002b).
Además, los cultivos donde las substancias mencionadas se hallaron, fueron fuertemente estimulados con phytohemagglutinin, IL-2, suero sanguíneo con anticuerpos anti interferón humano, y otros agentes (Papadopulos-Eleopulos et al 1996; de Harven 1998, 2003). Estos estimulantes de cultivo son agentes oxidantes y podía esperarse que estimulasen la expresión de retrovirus endógenos (Papadopulos-Eleopulos et al 1996). No existen en la literatura médica los controles requeridos para estos experimentos. Es interesante que ni “el VIH mismo” ni ningún marcador de VIH puede ser encontrado cuando los cultivos, supuestamente infectados, son tratados con antioxidantes (Papadopulos-Eleopulos 1988, 1998(9; Papadopulos-Eleopulos et al 1992, 1993).
Desgraciadamente, estas supuestas "proteínas del VIH" o "antígenos del VIH" son usados como antígenos en las pruebas serológicas para VIH, y esto explica la completa falta de especificidad de estas pruebas.
5. El llamado ARN del VIH no es un marcador específico del VIH.
LAS PRUEBAS PARA VIH NO PUEDEN DIAGNOSTICAR LA INFECCIN POR VIH
Además, la leyenda de los mono portadores del bichito y que contagiaron algunas personas es una mentira. Se plantaron el coctail a un mono en un laboratorio, y lo presentaron como un mono salvaje de la jungla. Gallo & co infectaron la comunidad de los gays en Nueva York con la enfermedad mediante un proyecto de vacunación contra hepatitis. Igualmente la población en Africa se contaminó mediante un programa de las banderillas contra la varicela. Intencionalmente. Otras enfermedades "raras" como
Nodding disease en Africa, son fruto del bioterrorismo practicado por los judíos infrahumanos, desafortunadamente en el poder en los EE.UU. Hablamos de Papercliff, de los jefes y los agentes de la CIA, Kissinger, Rockefellers, la familia Bush etc.
Kissinger fue detrás (con el judío Rockefeller, el propietario de Monsanto) de la fumigación masiva del agente Orange en Vietnam. Aún hoy en día hay niños vietnamitas que nacen deformados por el dicho producto químico.
 ienso:
ienso: