El ADN antiguo y la filogenia humana
Los estudios genéticos están iluminando el oscuro origen del sapiens gracias a que a partir de los diferentes alelos y haplotipos que portan los individuos actuales y antiguos, es posible estimar la fecha del ancestro común más reciente. La Teoría de la Coalescencia propone que, en una población dotada de una cierta variabilidad genética, y para genes neutros que no afecten a la viabilidad de un individuo, los diferentes alelos presentes en ella deberían haber tenido un sólo ancestro común, es decir, provenir de una única secuencia original.
Hace años se popularizó la llamada Eva mitocondrial (hipótesis Arca de Noé), o ancestro común más reciente para el ADN-mt. Sin embargo, los humanos actuales no provenimos de una sola mujer: Los modelos teóricos y computacionales muestran que número de individuos capaces de reproducirse fue siempre de al menos 4.000. Estos estudios cuentan con un problema importante: la calibración del reloj molecular, del número de sustituciones (mutaciones) por lugar del genoma (sitio) y unidad de tiempo.
La
paleogenética, en definitiva, ha aportado un grado muy elevado de certeza al conocimiento de:
La estructura formada por las diferentes poblaciones humanas en un momento determinado, incluyendo su tamaño.
La fecha de la separación entre poblaciones (pero DeSalle, 2016).
La movilidad de estas poblaciones y los eventos de hibridación.
La adscripción de un fósil a una determinada población, tras su análisis genético.
Carles Lalueza-Fox (2013) repasa algunos problemas de interpretación:
Los problemas asociados con los relojes moleculares.
La hipótesis de reloj molecular se basa en la regularidad del proceso de mutación en regiones genéticas neutrales a lo largo de tiempo, implicando así la posibilidad de utilizarlo como un estimador de tiempo para la evolución molecular. Hay, sin embargo, algunos problemas con la exactitud de un reloj molecular, de forma que en cada nueva toma de muestras a menudo se produce un recalibrado.
La diversidad genética actual (ya sea una población de estudio o de una especie) no está caracterizada en su totalidad. Una inclusión adicional en la muestra, puede atrasar las fechas considerablemente.
La tasa de mutación es una estimación, con pruebas contradictorias.
Hay que disponer de fechas exactas, pero el registro fósil no es muy preciso.
Hay que trabajar con regiones genómicas selectivamente neutras, pero la existencia de elementos reguladores ubicuos y barridos selectivos no detectados hace que su derterminación no sea obvia.
La diferencia entre la divergencia de secuencias genéticas y la divergencia de especies.
La teoría de la coalescencia nos permite ir hacia atrás en el tiempo a partir de la variación genética existente hasta encontrar antepasados comunes, lo que proporciona inferencias sobre la demografía de la población y la divergencia genética.
Los tiempos de coalescencia siempre son anteriores al de divergencia de las especies, debido a que en el momento de la separación existía una cierta variación genética. Estos tiempos serán más cercanos en la medida en que la población ancestral haya sido más pequeña y con ello más próxima genéticamente.
Las limitaciones de los marcadores uniparentales (ADN-mt y ADN-Y)
Los marcadores uniparentales han fracasado en la detección de los procesos evolutivos reales: Cuando el tamaño de la población permanece constante durante largo tiempo, el ADN uniparental tiende a aglutinarse en algún momento, de forma que los eventos genéticos anteriores son indetectables.
La forma en que la expresión de un genoma configura el fenotipo (incluyendo la morfología y la cognición).
Las publicaciones de los genomas humano y del chimpancé no cumplieron las expectativas para la comprensión de la base genética de las diferencias morfológicas (y cognitivas) que existen entre estas dos especies. El problema reside en las dificultades en la comprensión de la función del gen y también en la complejidad del genoma que funciona por encima del nivel del ADN. Existen muchos elementos reguladores que interactúan con redes de genes. Genomas similares o incluso idénticos podrían producir diferentes fenotipos como resultado de diferencias en la regulación de la transcripción de genes.
Las restricciones físicas y químicas pueden provocar que sólo pueda surgir en el tiempo un determinado conjunto de rasgos adaptativos y que rasgos similares aparezcan de forma independiente en diferentes linajes de homínidos. Diferentes bagajes genéticos pueden producir el mismo fenotipo.
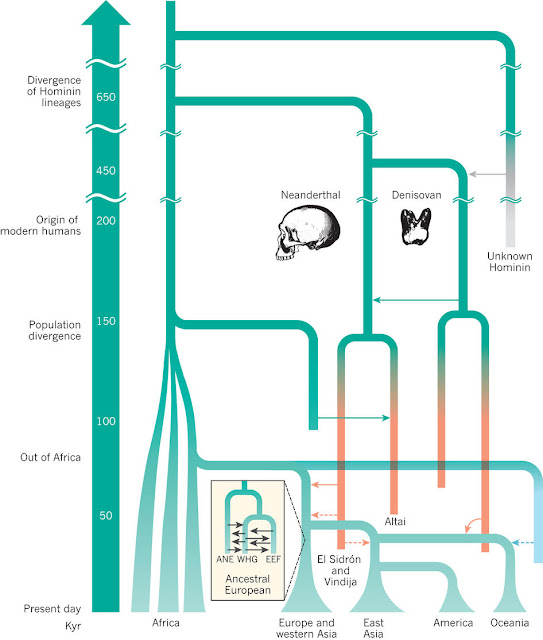
Relaciones entre poblaciones contemporáneas y momento aproximado de divergencia. Las líneas continuas y de puntos indican eventos de mezcla con pruebas más o menos sólidas respectivamente.
ANE: Euroasiáticos del Norte antiguos.
EEF: Agricultores tempranos europeos.
WHG: Cazadores recolectores europeos occidentales.
Nielsen et al, 2017.